Gene Drift - faktor evolucije: razlog, vrijednost, primjeri
Uz prirodnu selekciju postoji još jedan faktor koji može utjecati na povećanje sadržaja mutantnog gena. U nekim slučajevima, čak može pomaknuti normalan alel. Ovaj fenomen naziva se "pomicanje gena u populaciji". Razmotrimo detaljnije što je to proces i koje su njegove posljedice. 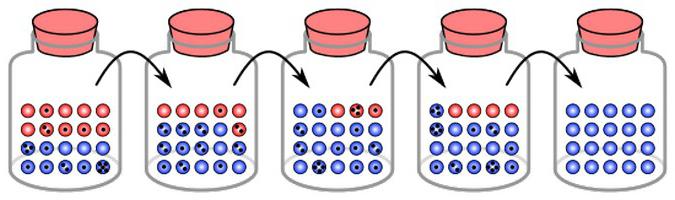
Opće informacije
Klizanje gena, primjeri koji će biti navedeni u članku u nastavku, određene su promjene koje se bilježe iz generacije u generaciju. Smatra se da ovaj fenomen ima svoje mehanizme. Neki istraživači su zabrinuti da u genskom fondu mnogih, ako ne i svih nacija, količina anomalnih gena koji se pojavljuju trenutno raste vrlo brzo. Utvrđuju nasljednu patologiju, stvaraju preduvjete za razvoj mnogih drugih bolesti. Također se vjeruje da je patomorfoza (promjena znakova) raznih bolesti, uključujući i one mentalne prirode, upravo pomak gena. Pojava o kojoj se radi pojavljuje se brzim tempom. Kao rezultat toga, broj mentalni poremećaji uzima nepoznate oblike, postaje neprepoznatljiv u usporedbi s njihovim opisom u klasičnim izdanjima. Istodobno, značajne promjene su uočene izravno u strukturi samog psihijatrijskog morbiditeta. Na primjer, pomicanje gena briše neke oblike shizofrenije koji su se susreli ranije. Umjesto toga, postoje patologije koje se teško mogu odrediti suvremenim klasifikatorima. 
Wrightova teorija
Slučajni pomak gena proučavan je pomoću matematičkih modela. Koristeći to načelo, Wright je izveo teoriju. Smatrao je da je presudna važnost pomaka gena pod konstantnim uvjetima zabilježena u malim skupinama. Oni postaju homozigotni i varijabilnost se smanjuje. Wright je također vjerovao da su, kao rezultat promjena u skupinama, mogle formirati negativne nasljedne osobine. Kao rezultat toga, cjelokupna populacija može umrijeti bez doprinosa razvoj vrste. Istovremeno, selekcija igra veliku ulogu u mnogim skupinama. U tom smislu, genetske varijacije unutar populacije opet će biti beznačajne. Postupno će se grupa dobro prilagoditi uvjetima okoliša. Međutim, naknadne evolucijske promjene ovisit će o pojavljivanju povoljnih mutacija. Ovi procesi su vrlo spori. U tom smislu, evolucija velikih populacija nije karakterizirana velikom brzinom. U skupinama srednjih vrijednosti uočava se povećana varijabilnost. Istodobno se slučajno stvara i stvaranje novih korisnih gena, što ubrzava evoluciju. 
Wrightovi zaključci
Kada se iz populacije izgubi jedan alel, može se pojaviti zbog određene mutacije. Ali ako je vrsta podijeljena u nekoliko skupina, u jednom od kojih nedostaje jedan element, u drugom, drugi, onda se gen može migrirati od mjesta gdje je, do mjesta gdje nije. Dakle, varijabilnost će ostati. S obzirom na to, Wright je zaključio da će se razvoj brže dogoditi u onim vrstama koje su podijeljene u brojne različite najveće populacije. Istodobno je između njih moguća neka migracija. Wright se složio s tim prirodna selekcija igra vrlo značajnu ulogu. Međutim, uz ovaj rezultat evolucije dolazi i do gena. Identificira trajne promjene unutar prikaza. Osim toga, Wright je vjerovao da su mnoge karakteristične značajke koje su nastale zbog zanošenja indiferentne, au nekim slučajevima čak i štetne za održivost organizama. 
Sporovi istraživača
Bilo je nekoliko mišljenja o Wrightovoj teoriji. Na primjer, Dobzhansky je vjerovao da je besmisleno postavljati pitanje koji je od važnijih čimbenika - prirodna selekcija ili genetski zanos. Objasnio je to svojom interakcijom. U biti, moguće su sljedeće situacije:
- Ako u razvoju jedne ili druge vrste selekcija zauzme vodeću poziciju, označit će se ili smjerna promjena frekvencija gena ili stabilno stanje. Potonji će biti određen uvjetima okoline.
- Ako je tijekom duljeg razdoblja pomak gena značajniji, tada usmjerene promjene neće biti uzrokovane prirodnim okolišem. Istodobno, nepovoljni znakovi, čak iu malim količinama, mogu se u grupi dovoljno proširiti.
Međutim, treba napomenuti da sam proces promjene, kao i uzrok pomicanja gena, danas nije dovoljno istražen. U tom smislu ne postoji jedinstveno i konkretno mišljenje o ovoj pojavi u znanosti.
Odvaganje gena je čimbenik evolucije
Zbog promjena, zabilježena je promjena frekvencija alela To će se dogoditi dok ne dostignu stanje ravnoteže. To jest, zanošenje gena je izolacija jednog elementa i fiksacija drugog. U različitim skupinama takve se promjene događaju neovisno. U tom smislu, rezultati genetskog drifta u različitim populacijama su različiti. Na kraju, jedan je skup elemenata fiksiran u nekim, drugi u drugima. Odvagnjavanje gena, s jedne strane, stoga dovodi do smanjenja raznolikosti. Međutim, u isto vrijeme, ona također uzrokuje razlike između skupina, razlike na nekim osnovama. To, pak, može poslužiti kao osnova za specijaciju. 
Omjer utjecaja
U procesu razvoja, genetski zanos surađuje s drugim čimbenicima. Prije svega, odnos se uspostavlja s prirodnom selekcijom. Omjer doprinosa ovih čimbenika ovisi o nizu okolnosti. Prije svega to je određeno intenzitetom selekcije. Druga okolnost je veličina skupine. Dakle, ako su intenzitet i snaga visoki, slučajni procesi imaju zanemariv učinak na dinamiku genetskih frekvencija. Istovremeno, u malim skupinama s neznatnim razlikama u sposobnosti, učinak promjena je neusporedivo veći. U takvim slučajevima može se fiksirati manje adaptivni alel, a više adaptivni će biti izgubljen.
Učinci promjene
Jedan od glavnih rezultata genetskog drifta je osiromašenje različitosti unutar grupe. To je zbog gubitka nekih alela i fiksacije drugih. Proces mutacije, naprotiv, pridonosi obogaćivanju genetske raznolikosti unutar populacija. Zbog mutacije, izgubljeni alel se može pojavljivati iznova i iznova. S obzirom na činjenicu da je genetski drift usmjeren proces, istodobno s smanjenjem intrapopulacijske raznolikosti, povećava se razlika između lokalnih skupina. Protivljenje ovom fenomenu je migracija. Dakle, ako je u jednoj populaciji alel "A" fiksiran, au drugom "a", onda se unutar tih grupa raznolikost ponovno pojavljuje. 
Konačni rezultat
Rezultat genetskog drifta bit će potpuna eliminacija jednog alela i konsolidacija drugog. Što se element češće pojavljuje u skupini, veća je vjerojatnost njegove fiksacije. Kao što neki izračuni pokazuju, mogućnost vezanja jednaka je učestalosti alela u populaciji.
mutacije
One se pojavljuju u prosjeku s učestalošću od 10-5 po genu po gametu po generaciji. Svi aleli koji se nalaze u skupinama jednom su se pojavili zbog mutacija. Što je populacija manja, to je manja vjerojatnost da će svaka generacija imati barem jednu osobu - nositelja nove mutacije. S brojem sto tisuća, svaka skupina potomaka s vjerojatnošću bliskom jedinstvu ima mutirani alel. Međutim, njegova učestalost u populaciji, kao i mogućnost njezina fiksiranja bit će prilično niska. Vjerojatnost da će se ista mutacija pojaviti u istoj generaciji, barem kod jednog pojedinca s populacijom od 10, je zanemariva. Međutim, ako se to dogodi u ovoj populaciji, učestalost mutantnog alela (1 u 20 alela), kao i šanse za njegovo fiksiranje, bit će relativno visoka. U velikim populacijama pojavljivanje novog elementa događa se relativno brzo. U isto vrijeme, njezino fiksiranje je sporo. Nasuprot tome, male populacije očekuju dugu mutaciju. No, nakon njegove pojave, pričvršćivanje brzo prolazi. Iz toga se može izvesti sljedeći zaključak: vjerojatnost fiksiranja neutralnih alela ovisi samo o učestalosti mutacije. Istodobno, veličina populacije ne utječe na taj proces.
Molekularni sat
Zbog činjenice da je učestalost pojavljivanja neutralnih mutacija u različitim vrstama približno jednaka, brzina fiksiranja također bi trebala biti približno jednaka. Iz toga slijedi da bi broj promjena akumuliranih u jednom genu trebao biti povezan s vremenom neovisne evolucije ovih vrsta. Drugim riječima, što je duže razdoblje od trenutka odvajanja dvije vrste s jednog predaka, to će više razlikovati mutacijske supstitucije. Ovaj princip je temelj metode molekularnog evolucijskog sata. Time se definira vrijeme koje je prošlo od trenutka kada su se prethodne generacije raznih sustavnih skupina počele razvijati neovisno, neovisno jedna o drugoj.
Polling i Cukurkend studija
Ova dva američka znanstvenika su otkrila da je broj razlika u sekvenci aminokiselina u citokromu i hemoglobinu u određenim vrstama sisavaca viši, ranije su se njihovi evolucijski putovi divergirali. Nakon toga, ovaj uzorak je potvrđen velikom količinom eksperimentalnih podataka. Materijal je uključivao na desetke različitih gena i nekoliko stotina vrsta životinja, mikroorganizama i biljaka. Pokazalo se da se tijek molekularnih sati izvodi konstantnom brzinom. Ovo otkriće, zapravo, potvrđuje teorija o kojoj se radi. Sat je kalibriran zasebno za svaki gen. To je zbog činjenice da je učestalost pojave neutralnih mutacija u njima različita. U tu svrhu se procjenjuje broj supstitucija akumuliranih u određenom genu u taksonomima. Njihovo vrijeme divergencije pouzdano se utvrđuje paleontološkim podacima. Nakon kalibriranja molekularnog sata, oni se mogu dalje koristiti. Osobito, uz njihovu pomoć lako je izmjeriti vrijeme tijekom kojeg je došlo do divergencije (divergencije) između različitih taksona. To je moguće čak i ako njihov zajednički predak još nije identificiran u fosilnim zapisima.