Što je kromatin? Funkcije kromatina
Genetski materijal eukariotskih organizama ima vrlo složenu organizaciju. DNA molekule, nalazi u jezgri stanice, dio su posebne multikomponentne tvari - kromatina.
Definicija pojma
Kromatin je materijal stanične jezgre koja sadrži nasljednu informaciju, koja je složeni funkcionalni DNA kompleks sa strukturnim proteinima i drugim elementima koji osiguravaju pakiranje, skladištenje i realizaciju kariotskog genoma. U pojednostavljenom tumačenju, to je tvar koja čini kromosome. Pojam dolazi od grčkog "kroma" - boje, boje.
Koncept je Fleming uveo još davne 1880. godine, ali još uvijek postoje rasprave o tome što je kromatin sa stajališta njegove biokemijske kompozicije. Nesigurnost se odnosi na mali dio komponenti koje nisu uključene u strukturiranje genetskih molekula (neki enzimi i ribonukleinske kiseline).
U elektronskoj fotografiji interfazne jezgre, kromatin se vizualizira kao brojne mrlje tamne tvari, koje mogu biti male i raspršene ili kombinirane u velike guste nakupine.
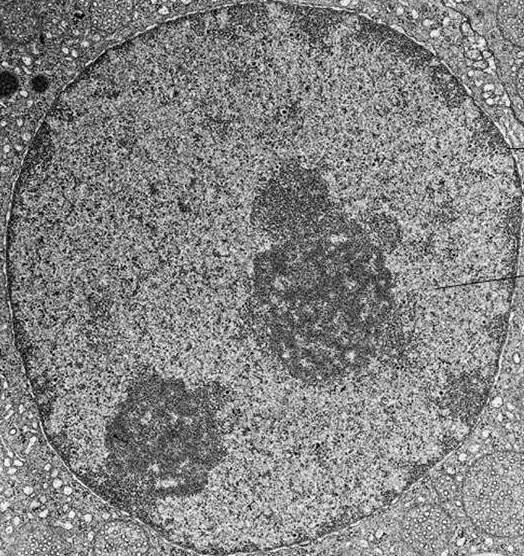
Kondenzacija kromatina tijekom stanične diobe dovodi do stvaranja kromosoma, koji su vidljivi čak iu konvencionalnom svjetlosnom mikroskopu.
Strukturne i funkcionalne komponente kromatina
Kako bi se utvrdilo što je kromatin na biokemijskoj razini, znanstvenici su ovu tvar izlučili iz stanica, prenijeli u otopinu i proučavali sastav i strukturu sastojaka u ovom obliku. Korištene su kemijske i fizikalne metode, uključujući tehnologije elektronske mikroskopije. Pokazalo se da je kemijski sastav kromatina za 40% predstavljen dugim molekulama DNA i gotovo 60% različitim proteinima. Potonji su podijeljeni u dvije skupine: histoni i ne-histoni.
Histoni su velika obitelj jezgrenih nuklearnih proteina koji se snažno vežu na DNA, tvoreći strukturni kostur kromatina. Njihov broj je približno jednak postotku genetskih molekula.
Ostatak (do 20%) proteinske frakcije pada na DNA-vežuće i prostorno-modificirajuće proteine, kao i enzime uključene u proces čitanja i kopiranja genetske informacije.
Osim glavnih elemenata, u sastavu kromatinskih ribonukleinskih kiselina (RNA), glikoproteini, ugljikohidrati i lipidi nalaze se u maloj količini, ali je pitanje njihove povezanosti s kompleksom DNA pakiranja još uvijek otvoreno.
Histoni i nukleosomi
Molekularna težina histona varira od 11 do 21 kDa. Veliki broj osnovnih aminokiselinskih lizina i ostataka arginina daje tim proteinima pozitivan naboj, što doprinosi stvaranju ionskih veza s suprotno nabijenim fosfatnim skupinama dvostruke spirale DNA.
Postoji 5 vrsta histona: H2A, H2B, H3, H4 i H1. Prve četiri vrste uključene su u formiranje glavne strukturne jedinice kromatina - nukleosoma, koji se sastoji od jezgre (proteinske jezgre) i DNA omotane oko nje.
Nukleosomska jezgra je predstavljena s osam molekula oktamernog kompleksa histona, koji uključuje tetramer H3-H4 i dimer H2A-H2B. Područje DNA s duljinom od oko 146 nukleotidnih parova je namotano na površinu proteinske čestice, formirajući 1,75 zavojnice, i prelazi u linkersku sekvencu (približno 60 bp) koja povezuje nukleosome jedan s drugim. H1 molekula veže se na linker DNA, štiteći je od djelovanja nukleaza.
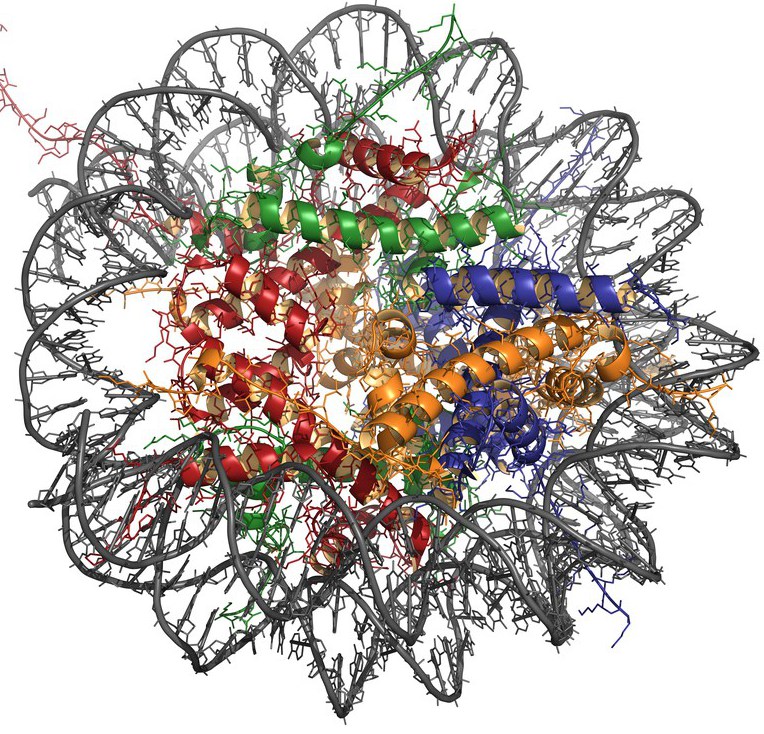
Histoni se mogu podvrgnuti različitim modifikacijama, kao što su acetilacija, metilacija, fosforilacija, ADP-ribozilacija i interakcija s ubikvitin proteinom. Ti procesi utječu na prostornu konfiguraciju i gustoću DNA pakiranja.
Ne-histonski proteini
Postoji nekoliko stotina različitih ne-histonskih proteina s različitim svojstvima i funkcijama. Njihova molekularna težina varira od 5 do 200 kDa. Posebna skupina sastoji se od specifičnih proteina, od kojih je svaki komplementaran specifičnoj regiji DNA. Ova grupa uključuje 2 obitelji:
- "cinkovi prsti" - prepoznaju fragmente od 5 parova nukleotida;
- homodimeri su karakterizirani strukturom helix-turn-helix u fragmentu povezanom s DNA.
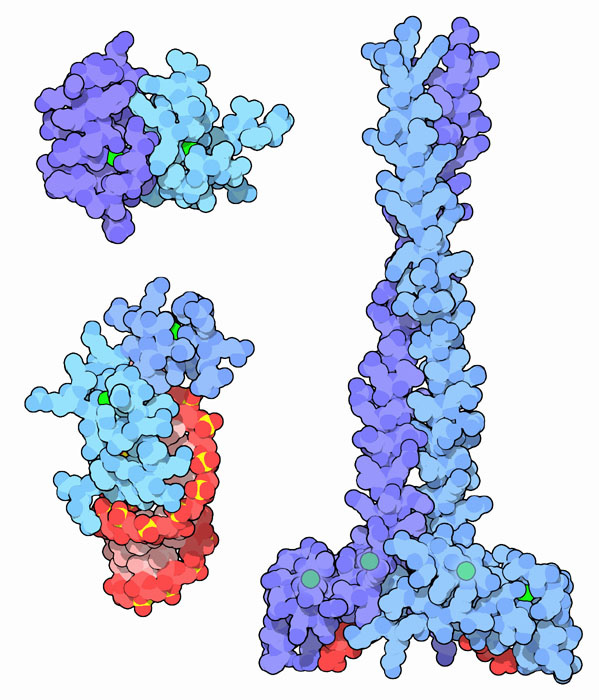
Najbolji su proučavani takozvani proteini visoke mobilnosti (proteini HGM) koji su trajno povezani s kromatinom. Ovo ime obitelji primljeno je zbog velike brzine kretanja molekula proteina u elektroforetskom gelu. Ova skupina zauzima većinu ne-histonske frakcije i uključuje četiri glavna tipa proteina HGM: HGM-1, HGM-14, HGM-17 i HMO-2. Oni obavljaju strukturne i regulatorne funkcije.
Ne-histonski proteini također uključuju enzime koji osiguravaju transkripciju (sinteza glasničke RNA), replikaciju (dupliranje DNA) i popravak (popravak oštećenja u genetskoj molekuli).
Razine zbijanja DNA
Osobitost strukture kromatina je takva da omogućuje da se DNA lanci ukupne duljine više od metra uklope u jezgru promjera oko 10 mikrona. To je moguće zahvaljujući višestupanjskom sustavu pakiranja genetskih molekula. Opća shema zbijanja uključuje pet razina:
- nukleosomni filament promjera 10-11 nm;
- fibril 25–30 nm;
- domene petlje (300 nm);
- Vlakno debljine 700 nm;
- kromosomi (1200 nm).
Ovaj oblik organizacije osigurava smanjenje duljine izvorne DNA molekule za 10 tisuća puta.
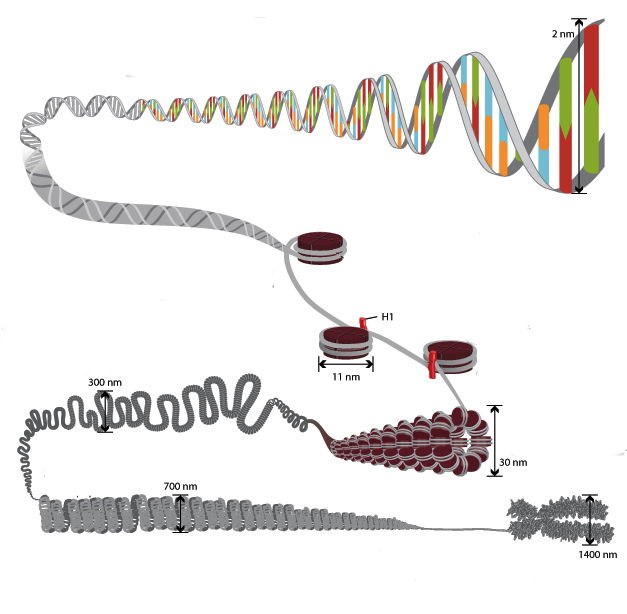
Nit od 11 nm formiran je nizom nukleosoma povezanih s DNA veznim regijama. Na elektronskim mikrografama takva struktura podsjeća na perle nanizane na liniji za ribolov. Nukleosomna vlakna se presavijaju u zavojnicu kao solenoid, tvoreći fibril debljine 30 nm. Histon H1 je uključen u njegovo formiranje.
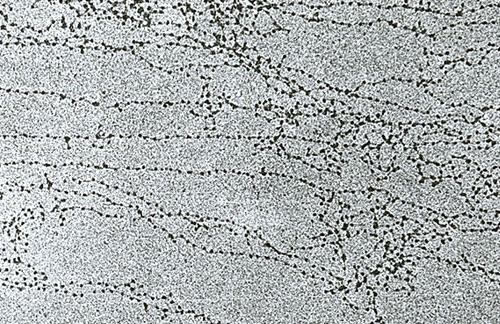
Solenoidna fibril se savija u petlje (tzv. Domene), koje su fiksirane na potpornu intranuklearnu matricu. Svaka domena sadrži od 30 do 100 tisuća parova nukleotida. Ova razina zbijanja karakteristična je za interfazni kromatin.
Struktura debljine 700 nm formirana je spiralizacijom fibrila domene i zove se kromatid. S druge strane, dvije kromatide čine petu razinu DNA organizacije - kromosom promjera 1400 nm, koji postaje vidljiv u fazi mitoze ili mejoze.
Dakle, kromatin i kromosom su oblici pakiranja genetskog materijala koji ovise o životnom ciklusu stanice.
kromosom
Kromosom se sastoji od dvije sestrinske kromatide identične jedna drugoj, od kojih je svaka formirana jednom superkovanom molekulom DNA. Polovice su povezane posebnim vlaknastim tijelom koje se naziva centromer. U isto vrijeme, ova struktura je sužavanje koje razdvaja svaku kromatidu na ramena.
Za razliku od kromatina, koji je strukturni materijal, kromosom je diskretna funkcionalna jedinica, karakterizirana ne samo strukturom i sastavom, već i jedinstvenim genetskim skupom, kao i određenom ulogom u provedbi mehanizama nasljednosti i varijabilnosti na staničnoj razini.
Euchromatin i heterochromatin
Kromatin u jezgri postoji u dva oblika: manje spiraliziran (euchromatin) i kompaktniji (heterochromatin). Prvi oblik odgovara transkripcijski aktivnim dijelovima DNA i stoga nije tako čvrsto strukturiran. Heterokromatin se dijeli na opcionalno (može se kretati od aktivnog do gustog neaktivnog oblika ovisno o stupnju životnog ciklusa stanice i potrebi primjene određenih gena) i konstitutivnim (stalno kondenziranim). Tijekom mitotske ili meiotičke podjele, svi kromatini su neaktivni.
Konstitutivni heterohromatin pronađen je u blizini centromera iu terminalnim dijelovima kromosoma. Rezultati elektronske mikroskopije pokazuju da takav kromatin zadržava visok stupanj kondenzacije ne samo u fazi stanične diobe, već i tijekom interfaze.
Biološka uloga kromatina
Glavna funkcija kromatina je gusto ispunjena velikom količinom genetskog materijala. Međutim, jednostavno stavljanje DNA u jezgru nije dovoljno da bi stanica funkcionirala. Potrebno je da te molekule ispravno "rade", tj. Mogu prenijeti informacije sadržane u njima kroz sustav DNA-RNA-protein. Osim toga, stanica treba distribuirati genetski materijal tijekom podjele.
Uređaj za kromatin potpuno ispunjava ove ciljeve. Proteinski dio sadrži sve potrebne enzime, a osobitosti strukture omogućuju im interakciju s određenim dijelovima DNA. Stoga je druga važna funkcija kromatina da osigura sve procese povezane s provedbom nuklearnog genoma.